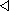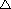Zovšeobecnenie metódy BP na RC NN bolo prvýkrát prezentované v [16].
Logika celého procesu BP je veľmi podobná ako pre FF NN. Pri RC NN nás však
do veľkej miery zaujíma dynamika činnosti RC NN z dôvodu existencie
rekurentných synapsii. Teda už pri šírení signálu sa musia vytvárať podmienky
GS a až potom sa môže šíriť späť samotná chyba. Pod dynamikou NN systému budeme
rozumieť nejakú funkciu ,
kde
je stav neurónu v čase ,
teda
(2.26)
Teda v prípade metódy BP na RC NN musíme hľadať také adaptačné pravidlo zmeny
SV, ktoré :
bude minimalizovať chybu v zmysle BP prístupu
bude mať podmienky na dosiahnutie stavu stability v bode
kedy je
.
Teda hlavným rozdielom medzi BP na RC NN a FF NN je
problém neurodynamiky. RC NN musí vykazovať stabilné vlastnosti
pri šírení signálu dopredu ako aj pri spätnom šírení
chyby. Inak môžu vzniknúť nekontrolovateľné procesy a
celý systém bude nefunkčný. Je zrejmé, že exaktný popis modelu
neurónu a jeho dynamiky bude mať význam pri štúdiu dynamiky
celého systému. V literatúre tykajúcej sa NN sa používa tzv.
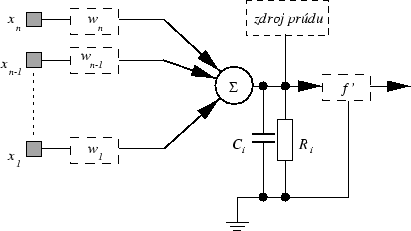
Ressistance-Capacity model neurónu ako
východisko pre popis RC NN viď obrázok 2.3.
Obrázok 2.3:
Ressistance-Capacity model neurónu
Ak tento model budeme považovať za jednoduchý elektrický obvod, potom v bode
V bude platiť Kirchhoffov zákon o súčte prúdov. Tento model má aj svoju
analógiu v biologických systémoch, kde
vodivosť medzi neurónmi "i" a "j"
priemerný potenciál neurónov na základe vstupu a výstupu
z neurónov
odpor neurónovej membrány neurónu "i"
kapacita neurónovej membrány neurónu "i"
je prahová funkcia neurónu "i"
Teda potom podľa Kirchoffovho zákona dostaneme :
(2.27)
kde
.
Táto rovnica vlastne determinuje podmienky rovnováhy v neuróne,
t.j.
na uvedenom modeli neurónu. Rovnicu (2.27)
je možné upraviť do tvaru (využijeme fyzikálne zákony)
(2.28)
kde
je funkcia .
Ešte raz je potrebné zdôrazniť, že (2.28)
predstavuje podmienky takej zmeny stavu neurónu ,
aby
došlo k GS. Riešením tejto diferenciálnej rovnice je bod
stability
v stavovom priestore.
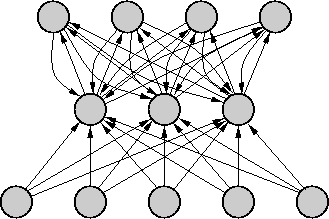
Obrázok 2.4:
Topológia RC NN
Ak sa teraz vrátime k reálnej topológii RC NN na obr. 2.4, tak celkovo
môžeme vyjadriť vstup do ľubovoľného neurónu ako
(2.29)
kde
je vstup do neurónu z externého sveta (,
samotný vstup).
Ďalej môžeme definovať chybový rozdiel ( pri predpoklade identickej výstupnej
funkcie
) ako
(2.30)
kde
je spomínaný stav výstupného neurónu "i" pri dosiahnutí
rovnovážneho stavu a
je očakávaná hodnota
neurónu v zmysle kontrolovaného učenia. Teda, ako u klasickej
metódy BP deklarujeme chybovú funkciu v tvare
(2.31)
kde k sa mení od 1 po počet neurónov vo výstupnej vrstve. V
konečnom dôsledku dostaneme všeobecný výraz pre adaptačné
pravidlo pre spojenie medzi ľubovolnými "s" a "r" neurónmi: